 |
|
| biologia |
|
|||||
|
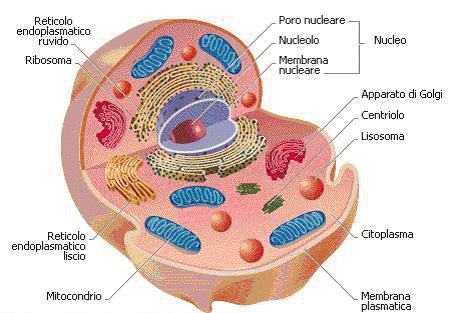
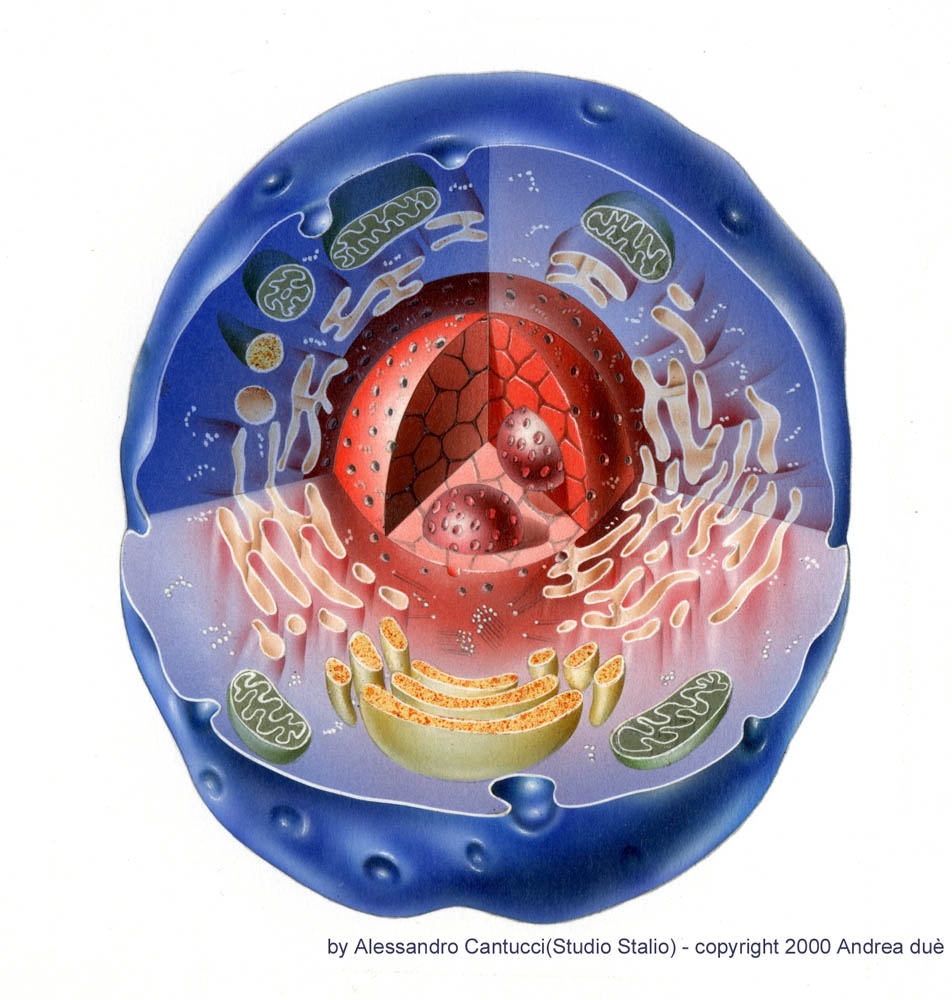
CELLULA
Le cellule sono i mattoni che costituiscono il corpo degli esseri viventi.
Ogni cellula è separata dall'ambiente circostante per mezzo di una membrana plasmatici, e nella cellula si possono distinguere 2 timenti fondamentali: il nucleo e il citoplasma.
Il citoplasma può essere diviso ulteriormente in una parte fluida il citosol (proteine, ioni zuccheri e altre molecole solubili) e formazioni definite organuli che possono essere delimitati o meno da membrane.
MEMBRANA PLASMATICA
È composta da fosfolipidi, che ne costituiscono il 20/30 %, da colesterolo e proteine 50/60 %, glicolipidi e glicoproteine.
Se osservata al microscopio elettronico, appare con una struttura trilaminare, con due strati elettrodensi, separati da uno strato chiaro.
Gli strati elettrodensi sono formati dalla porzione idrofilica (teste polari) di molecole fosfolipidiche, mentre le porzioni idrofobiche di quest'ultima sono rivolte all'interno, e insieme a molecole di colesterolo costituiscono lo strato centrale. Il colesterolo limita i movimenti laterali dei fosfolipidi e diminuisce la fluidità della membrana, stabilizzandola.
Sul versante sia interno che esterno della membrana di inseriscono le proteine estrinseche. ½ sono poi proteine integrali, idrofobiche incorporate saldamente fra i vari strati della membrana, alcune proteine integrali si estendono x l'intero spessore della membrana e vengono perciò dette TRANSMEMBRANA.
Si può considerare la membrana plasmatici come un mosaico di proteine immerse in un doppio strato lipidico fluido.
Le proteine transmembrana costituiscono canali che si aprono e si chiudono per lasciar passare dall'esterno all'interno e viceversa non solo ioni e piccole molecole solubili, ma anche acqua (tubulo prossimale del rene).
Vi sono proteine transmembrana che fungono da recettori, cioè hanno la capacità di legare ormoni chiamati neurotrasmettitori.
Possiamo affermare che le principali funzioni della membrana cellulare riguardano l'isolamento della cellula dall'ambiente esterno, regolazione degli scambi metabolici, e il supporto per la formazione dei tessuti.
La membrana plasmatici è permeabile ad acqua, ossigeno, anidride carbonica, piccole molecole idrofobiche (etanolo), ormoni stereoidei, ma è impermeabile a ioni come Na+, k+, per tanto costituisce un'efficace barriera dall'ambiente esterno.
Il passaggio di molecole attraverso le membrane può essere attivo (cioè richiede una sorgente energetica ATP) oppure passivo (cioè avviene senza dispendio di energia).
Gli scambi passivi possono avvenire in 4 modi:
Gli scambi attivi avvengono in 2 modi:
Di distinguono 3 tipi di endocitosi:
pinocitosi: le vescicole si riempiono di liquido extracellulare contenente lipidi, zuccheri o amminoacidi;
endocitosi:: è mediata da recettori, le vescicole contengono un alta concentrazione di una specifica molecola detta ligando (rappresentata da ormoni, fattori della crescita) che si è legata a recettori presenti sulla membrana plasmatici.
fagocitosi: le vescicole contengono particelle solide che possono essere molto grandi; le vescicole possono successivamente fondersi con i lisosomi x digerire le particelle fagocitate (funzione tipica dei globuli bianche e dei granulociti)
CITOPLASMA E ORGANULI
La parte fluida del citoplasma è detta Citosol, nel quale si ha una concentrazione relativamente alta di proteine, molte delle quali sono enzimi.
Il citosol contiene anche zuccheri spesso sotto forma di glicogeno( come nel fegato), amminoacidi e gocciole lipidiche.
Il citosol in definitiva è un sistema celloidale polifasico con una fase disperdente costituita da acqua ioni e micromolecole e una fase dispersa rappresentata da macromolecole che modificano la viscosità.
Gli organuli presenti nel citoplasma sono rappresentati fondamentalmente da: ribosomi, mitocondri, lisosomi,perossisomi.
RIBOSOMI
Sono piccoli granuli privi di membrana composti dal 60% di RNA e il restante 40% da proteine.
Possono essere dispersi nel citoplasma oppure fissati ad una parte del reticolo endoplasmatico, per questo motivo viene detto rugoso(RER). Sono deputati alla sintesi proteica; le proteine sintetizzate vengono riversate nel RER e poi inviate all'apparato di Golgi che provvede a smistarle verso i lisosomi, la membrana plasmatica ecc.
Tali proteine presentano nel loro tratto iniziale una speciale sequenza amminoacidica detta sequenza segnale, che viene rimossa appena la proteina penetra nel RER.
Le proteine prodotte nei ribosomi liberi permangono nel citosol.
MITOCONDRI
Sono organuli di forma sferica, rivestiti da membrana, deputati a fornire alle cellule l'ATP necessario x molteplici funzioni.
Si ritiene che i mitocondri siano stati microrganismi simili ai batteri, successivamente evolutosi, in effetti contengono un proprio DNA, che si replica in modo indipendente rispetto a quello del nucleo e sintetizzano in proprio numerose proteine.
Ogni mitocondrio possiede due membrane una esterna e una interna che definiscono 2 spazi: lo spazio intermembranoso e lo spazio matrice.
La membrana esterna contiene una proteina chiamata porina che svolge un ruolo chiave x il controllo della permeabilità; la membrana interna è altamente impermeabile ai piccoli ioni a causa del suo alto contenuto di fosfolipidi.
La membrana interna presenta numerosi ripiegamenti dette creste, che servono x aumentare la superficie è la sede degli enzimi della catena respiratoria nonché della ATPsintassi responsabile della produzione di energia.
Lo spazio intermembranoso contiene molecole che diffondono attraverso la membrana esterna, ATP e ioni che vengono pompati al di fuori della matrice durante la fosforilazione ossidativa.
Lo spazio matrice contiene enzimi per l'ossidazione degli acidi grassi, e dell'acido piruvico, nonché dell'acido citrico. Esso è anche la sede del DNA mitocondriale e della sua duplicazione.
I mitocondri sono molto numerosi nelle cellule che consumano molta energia e quindi richiedono grosse quantità di ATP (tipo il tessuto muscolare cardiaco).
LISOSOMI
Sono organuli circondati da membrana, essi funzionano da apparato digerente intracellulare, destinato a degradare materiale captato x endocitosi o fagocitosi.
I lisosomi provengono dall'apparato di Golgi e appaiono come vescicole.
PEROSSISOMI
Sono organuli circondati da membrane, contenenti enzimi coinvolti nell'ossidazione, in particolare di acidi grassi a catene lunghe, inoltre contengono l'enzima per ossidasi che trasforma l'acqua ossigenata (una molecola molto dannosa per la cellula) in acqua.(le cellula apatiche ne contengono molto)
MEMBRANE INTRACELLULARI
Comprendono il reticolo endoplasmatico liscio, il reticolo endoplasmatico rugoso, l'apparato di Golgi e numerose vescicole che derivano dal reticolo endoplasmatico e dall'apparato di Golgi.
RETICOLO ENDOPLASMATICO
Il reticolo endoplasmatico (RE) è un intreccio involuto e tridimensionale di spazi delimitati da una membrana, che si estende attraverso il citoplasma e genera un timento subcellulare ben definito e separate dal citoplasma. Questo timento ha molte ramificazioni appiattite (le cisterne) continue tra loro e in contatto con l'involucro nucleare. Il legame di migliaia di ribosomi (di solito nella regione delle grandi cisterne) conferisce alla superficie del reticolo un aspetto granuloso da cui deriva il nome di reticolo endoplasmatico ruvido. In altre regioni della cellula il reticolo è privo di ribosomi. Questa forma, che prende il nome di reticolo endoplasmatico liscio, è continua con il reticolo ruvido, ed è la sede della biosintesi dei lipidi e di una varietà di altri processi importanti,compreso il metabolismo di alcuni farmaci e di composti tossici.
Le proteine e ilipidi sintetizzati dai ribosomi nel reticolo rugoso attraversano uno speciale reticolo endoplasmatico di transazione, da qui vengono avvolti in vescicole di trasporto e veicolate all'apparato di Golgi.
APPARATO DI GOLGI
Quasi tutte le cellule eucariotiche possiedono l'apparato (o complesso) di Golgi, sacche membranose (cisterne) disposte in strati appiattiti. L'apparato di Golgi prende il nome dal suo scopritore, Camillo Golgi. Ha 3 funzioni principali;
CITOSCHELETRO
È una impalcatura proteica intracellulare che conferisce al citoplasma flessibilità e resistenza. È formato di 3 componenti:
I microfilamenti sono composti principalmente da actina, sono sparsi nel citoplasma e costituiscono una rete al di sotto della membrana cellulare.
I filamenti intermedi stabilizzano la posizione degli organuli citoplasmatici;
E i microtubuli, sono strutture tubulari cave, costituite da tubuline.
Durante la mitosi formano il fuso mitotico,che distribuisce i cromosomi alle estremità opposte delle cellule.
Inoltre sono i principali componenti dei Cetrioli, delle ciglia e dei flagelli.
I cetrioli sono composti da 3 triplette di microtubuli, essi svolgono un ruolo chiave nella formazione del fuso mitotico.
Le ciglia sono estroflessioni cilindriche allungate della membrana cellulare, oscillando permettono il movimento di fluidi a secrezioni presenti sulla superficie della cellula (tipo l'epitelio che riveste le vie respiratorie).
Ed in fine i flagelli che assomigliano alle ciglia, ma sono più lunghi ,sono presenti esclusivamente negli spermatozoi e hanno la funzione di muovere le cellule nel fluido in cui sono contenute.
NUCLEO
È il più grande timento intracellulare, circondato da membrana, contiene il patrimonio genetico sotto forma di DNA. La maggior parte delle cellule hanno un solo nucleo, ci sono comunque cellule binucleate (come gli epatociti), e polinucleate (fibre muscolari) mentre altre cellula non hanno nucleo (eritrociti).
Il nucleo è il centro di controllo, che programma e coordina le varie attività della cellula.
È formato da 3 componenti:
la membrana nucleare separa il nucleo dal citoplasma ed è formata da due membrane ciascuna perforata da sottili pori, che regolano il passaggio delle molecole. La materia liquida interna al nucleo è il plasma nucleare, composto da una soluzione acquosa contenente, ioni, enzimi, ecc.
la cromatina è una sostanza granulare composta da lunghe molecole di DNA associate a proteine ISTONI (collana di perle). Durante la divisione cellulare la cromatina si addensa e forma delle strutture chiamate cromosomi.
I nucleoli sono corpuscoli che sintetizzano l'RNA ribosomiale. Questo tipo di RNA si lega a determinate proteine prodotte nel citoplasma per formare le due subunità costitutive dei ribosomi.
Il nucleo è circondato da un involucro a doppia membrana, ognuna delle quali ha funzioni differenti. le due membrane dell'involucro nucleare delimitano lo spazio nucleare che è continuo con il R.E.R.; presentano dei pori che stabiliscono una comunicazione fra il citosol e l'interno del nucleo.
NUCLEOTIDI E ACIDI NUCLEICI
I nucleotidi svolgono nella cellula numerose funzioni importatni. Essi trasportano l'informazione genetica, sono i principali trasportatori dell'energia chimica intracellulare e sono anche componenti strutturali di molti cofattori e secondi messaggeri.
I nucleotidi sono formati da 3 componenti caratteristici:
La molecola senza gruppo fosforico è detta nucleoside.
Le basi azotate sono derivati da due componenti: la purina e la pirimidina
Sia il DNA che l'RNA contengono 2 basi puriniche principali (A)l'adenina e la (G)Guanina e 2 basi pirimidiniche principali.
Sia nel DNA sia nell'RNA una delle basi
pirimidiniche è la (C)citosina,
mentre la seconda base pirimidinica è la (T)Timina nel DNA, e (U)l'uracile
nell'RNA. Raramente si trova
Gli acidi
nucleici sono polimeri di nucleotidi
in cui le varie subunità sono tenute unite da ponti fosfodiestere tra il gruppo
ossidrilico
Vi sono due tipi di acidi nucleici:l'RNA e il DNA.
I nucleotidi che fanno parte dell'RNA contengono Ribosio e le basi pirimidiniche sono l'uracile e la citosina.
Nel DNA,
i nucleotidi contengono 2'-deossiribosio
e le basi pirimidiniche sono
Il modello del DNA nativo è costituito da due catene antiparallele avvolte in una doppia elica destrorsa.
Le coppie di basi complementari A=T e G≡C* si formano all'interno dell'elica e sono tenute unite da legami idrogeno, mentre lo scheletro covalente idrofilico composto da zucchero e fosfato è all'esterno dell'elica.
Come suggerito dal modello di WATTON e CRICK e dalle regole di CHARGAFF, una base A è sempre appaiata con T, mentre G è appaiata con C.La struttura a doppia elica consente solo questi appaiamenti.
Vi sono diverse forme di struttura del DNA:
la forma B che è la struttura proposta da Watson e Crick è quella più stabile per una molecola di DNA.
Poi abbiamo la forma A e Z, la forma A è favorita in molte soluzioni relativamente povere di acqua. Il DNA è sempre organizzato in una doppia elica destrorsa, ma l'elica è un po' più larga e il numero di coppie di basi per ogni giro di elica e 11 invece di 10,5 del DNA di forma B.
La forma Z del DNA invece è radicalmente diversa dalla forma B; la differenza più evidente è la rotazione dell'elica in senso sinistrorso. ½ sono 12 coppie di basi per ogni giro di elica e la struttura appare più sottile e allungata. Lo scheletro covalente assume un andamento a zig zag.
Un tipo di sequenza comune nel DNA è quella chiamata palindromo. Termine che viene applicato alle regioni di DNA in cui vi sono ripetuti invertiti di una sequenza di basi con una doppia simmetria presente nelle due catene del DNA.
Quando la sequenza è presente in ciascuna catena del DNA, la sequenza viene detta ripetuto speculare.

ESEMPIO DI DNA PALINDROMO

ESEMPIO DI DNA RIPETUTO SPECULARE
RNA
L'RNA messaggero è il veicolo con cui l'informazione genetica viene trasferita dal DNA ai ribosomi per la sintesi delle proteine.
Se trasporta il codice per un solo polipeptide,l'mRNA è monocistronico, se codifica 2 o più polipeptidi diversi, l'mRNA è policistronico.
Negli eucarioti la maggior parte degli mRNA sono monocistronici.
L'RNA messaggero è soltanto una delle diverse classi di RNA presenti nella cellula.
Gli RNA di traferimento (RNA transfer) servono come molecole di adattamento nella sintesi delle proteine; essi dopo aver legato a una delle loro estremità un amminoacido, si appaiano all'mRNA in modo tale che l'AA venga inserito nella sequenza corretta. Gli RNA ribosomiali sono componenti strutturali dei ribosomi.
REPLICAZIONE DNA
La replicazione del DNA è una reazione di polimerizzazione che ha come reagenti i quattro tipi di desossiribonucleosidi trifosfati (dNTP: dATP, dCTP, dGTP, dTTP). Benché nel filamento venga incorporato solo un fosfato, i nucleotidi di partenza devono essere trifosfati, solo così posseggono infatti l'energia necessaria per la reazione. È necessaria la presenza di un DNA a filamento singolo che funge da stampo, che determina la sequenza del filamento da costruire.
La reazione è catalizzata dalle DNA polimerasi, enzimi capaci di costruire una nuova catena nel verso 5'-3' individuati da Arthur Kornberg nel 1958 tramite un famoso esperimento [1]. Esse non sono in grado di iniziare un filamento ex novo, possono solo allungare un filamento polinucleotidico preesistente. È necessario quindi un innesco. Questo consiste di solito in un breve frammento di RNA appaiato allo stampo, prodotto da una RNA polimerasi detta primasi.
Per iniziare la replicazione, il DNA a doppia elica deve essere parzialmente denaturato da particolari proteine. Queste sono le elicasi, enzimi che separano attivamente i due filamenti usando l'energia dell'ATP, e le proteine denaturanti, o proteine destabilizzatrici dell'elica, non enzimatiche, che possono denaturare il DNA legandosi selettivamente alle porzioni a singolo filamento e stabilizzandole. Queste attività producono una forca replicativa, che migra esponendo progressivamente filamenti non appaiati, che possono essere replicati.

![]()
Forca replicativa
Poiché le polimerasi lavorano solo in senso 5'-3', un filamento (chiamato Filamento a replicazione progressiva) può essere replicato in modo quasi continuo, man mano che viene esposto, l'altro (Filamento a replicazione regressiva) risulta disseminato da brevi filamenti di DNA di nuova sintesi (i frammenti di Okazaki), ognuno dei quali reca all'inizio l'innesco di RNA. I nuovi filamenti devono essere quindi completati mediante la rimozione degli inneschi da parte di endonucleasi e il riempimento degli spazi rimasti ad opera di polimerasi di riparazione. Successivamente tutti questi frammenti di DNA di nuova sintesi del filamento in ritardo vengono legati dalla DNA-ligasi.
Il risultato della replicazione sono due doppie eliche identiche (salvo errori avvenuti durante il processo, che portano alla sa di mutazioni) costituite da un filamento preesistente e uno neoformato: per questa ragione la replicazione si dice semiconservativa.
Nelle molecole di DNA circolari dei Procarioti si ha una sola regione di Origine della replicazione dal quale partono due forche replicative (la struttura prende il nome di bolla di replicazione). Quando le due forche si incontrano dal lato opposto la replicazione è completata.
Negli Eucarioti la replicazione di ogni cromosoma inizia in più punti
Un filamento di DNA, detto filamento
leading (a replicazione progressiva), viene sintetizzato in modo continuo;
l'altro, detto filamento lagging (o filamento a replicazione
regressiva), è dapprima formato da corti frammenti di DNA (i frammenti
di Okazaki) di 1-3 kilobasi. In seguito i frammenti sono uniti dall'enzima DNA
ligasi.
La sintesi di DNA non può iniziare ex novo, quindi l'enzima primasi
sintetizza corti inneschi di RNA complementari al DNA stampo. Nei procarioti
gli inneschi dei frammenti di Okazaki sono rimossi dall'enzima RNasi H e
dalla Polimerasi I. Negli Eucarioti altri enzimi rimuovono i primer e
Le DNA Polimerasi, affinché il processo di
replicazione sia efficace, necessitano di proteine che aumentino la loro
attività e le stabilizzino sul filamento. Le clamp-loading legano il DNA
alla giunzione tra l'innesco e lo stampo, le sliding-clamp si
congiungono a queste ultime, caricano
Lo svolgimento del DNA parentale è catalizzato
dall'enzima elicasi, che denatura il filamento sfruttando l'idrolisi
dell'ATP. Proteine che si legano al DNA a singolo filamento stabilizzano il DNA
denaturato in modo che
Per evitare che i filamenti si attorciglino le topoisomerasi introducono tagli
singoli (nel caso delle Topoisomerasi I) o doppi (nel caso delle Topoisomerasi
II). Queste rotture reversibili fungono da perni, che consentono al DNA di
ruotare liberamente.
La replicazione inizia in corrispondenza di
una specifica sequenza chiamata origine di replicazione, che costituisce
un sito di legame per proteine iniziatrici. Una proteina si lega all'origine e
inizia a svolgere il DNA; recluta in seguito altre proteine e, insieme a esse e
all'elicasi, continua la denaturazione. Le primasi inizia la sintesi dei
primer. La regione in cui i filamenti parentali si separano e i due nuovi
filamenti si allungano è denominata forca replicativa. Nei Procarioti,
il cui DNA è generalmente circolare, sono presenti due forche replicative che
dipartono dalla medesima origine. Quando le due forche si incontrano la
replicazione è terminata.
I genomi eucariotici sono spesso assai più grandi di quelli procariotici e
necessitano di origini multiple. Da ogni origine di replicazione nasce una bolla
replicativa che si espande in direzioni opposte. Due bolle replicative
entrate in contatto si fondono formandone una sola.
TRADUZIONE DEL DNA O SINTESI PROTEICA
La sintesi proteica è detta traduzione ed è il trasferimento di informazioni da un linguaggio (acidi nucleici) a un altro (amminoacidi). La traduzione si svolge in tre fasi: inizio,allungamento e terminazione.
La prima fase, l'inizio, comincia quando la
subunità ribosomiale più piccola si attacca al filamento di mRNA presso
l'estremità
La combinazione fra la subunità ribosomiale più piccola,l'mRNA e tRNA d'inizio è detta complesso d'inizio.Una volta che questo si è formato, la subunità ribosomiale più grossa si attacca a quella più piccola e il tRNA d'inizio di incastra nel sito P(peptide) della subunità più grossa, che è uno dei due siti di legame con tRNA.
All'inizio della seconda fase, quella di allungamento, il secondo codone dell'mRNA si colloca di fronte al sito A. un tRNA con un anticodone complementare al secondo codone di mRNA si incastra sulla molecola di mRNA e, con il suo amminoacido, viene a occupare il sito A del ribosoma. Una volta occupati entrambi i siti, si forma un legame peptidico tra i due amminoacidi, attaccando il primo al secondo.
Il primo tRNA viene liberato. Il ribosoma poi sposta avanti un codone della catena di mRNA e di conseguenza il secondo tRNA
ALTRA VERSIONE DELLA TRADUZIONE
La sintesi proteica (nota anche come traduzione genetica) costituisce la seconda fase del processo di espressione genica, ovvero il processo nel quale l'informazione contenuta nel DNA dei geni strutturali viene trasformata in proteine che vanno a formare ed a far vivere una cellula.
Nella sintesi proteica un filamento di RNA messaggero, complementare ad una data regione del DNA, è usato come stampo per la produzione di una specifica proteina. La relazione tra triplette di basi dell'RNA e gli amminoacidi delle proteine è ciò che chiamiamo codice genetico.
La sintesi procede in tre fasi: inizio, crescita e termine del polipeptide.
L'inizio della sintesi vede i ribosomi legarsi al
codone di avvio (start) dell'mRNA, che indica il punto in cui l'mRNA
comincia a codificare la proteina. Questo codone è generalmente AUG (adenina-uracile-guanina),
ma codoni diversi sono frequenti nei procarioti. Negli eucarioti e negli archeobatteri
l'amminoacido corrispondente al codone di avvio è la metionina. Nei batteri
invece la proteina inizia con
Il tRNA iniziatore, sia che rechi metionina o N-formil-metionina, accoppia le sue basi con quelle del codone di avvio e si lega al sito P del ribosoma. La sub-unità maggiore forma quindi un complesso con quella minore. A questo punto avviene la crescita. Un nuovo tRNA entra sul sito A del ribosoma ed accoppia le sue basi con quelle dell'mRNA. L'enzima peptidil transferasi crea un legame peptidico tra gli amminoacidi vicini. Appena questo accade, l'amminoacido sul sito P si stacca dal suo tRNA e si lega al tRNA sul sito A. Il ribosoma quindi si muove lungo l'mRNA spostando il tRNA dal sito A al sito P liberando nel contempo il tRNA vuoto. Questo processo è noto come traslocazione.
Questo processo continua finché il ribosoma non incontra uno dei tre possibili codoni di arresto (stop), dove avviene il termine. La crescita della proteina si interrompe ed i fattori di rilascio, proteine che simulano l'azione del tRNA, si legano al sito A e liberano la proteina nel citoplasma.
La sintesi delle proteine può avvenire molto rapidamente. Questo avviene perché più ribosomi possono legarsi ad un filamento di mRNA consentendo quindi lacostruzione simultanea di più proteine. Un filamento di mRNA con più ribosomi è chiamata polisoma.
Infine, dato che i procarioti non hanno nucleo, un filamento di mRNA può essere tradotto in proteina mentre viene creato per trascrizione dal DNA. Questo non è possibile negli eucarioti, in cui la traduzione avviene nel citoplasma mentre la trascrizione (biologia) avviene nel nucleo cellulare.
È possibile bloccare specificamente la sintesi proteica facendo usare inibitori specifici quali l'anisomicina e la cicloesimmide. La traduzione può anche essere bloccata per effetto di mutazioni genetiche come ad esempio le mutazioni con slittamento di fase (frame shift) le quali, possono essere ottenute con la delezione (o l'inserzione) di un singolo paio di basi.
TRASCRIZIONE
La trascrizione è un processo mediante il quale le informazioni contenute nel DNA vengono trascritte, per complementarietà delle basi, nell'acido ribonucleico (RNA). Essa richiede l'uso di particolari enzimi, RNA polimerasi DNA-dipendenti. Tali enzimi utilizzano nucleosidi trifosfati (nucleotidi con tre gruppi fosfato) per la formazione dell' RNA. Dai nucleosidi trifosfato verranno rimossi due gruppi fosfato per la formazione di un legame covalente (Un legame covalente si instaura quando una coppia di elettroni viene messa in comune fra due atomi.)tra un nucleotide e il seguente. Si tratta di una reazione di condensazione.
L'RNA polimerasi si legherà solo in particolari sequenze, promotori, che non sono trascritte. Dal promotore inizieranno ad inserirsi i nucleosidi trifosfato per formare una sequenza di nucleotidi che sarà complementare al filamento di DNA in questione. Dopo l'individuazione del promotore, l'RNA polimerasi srotolerà il DNA a doppia elica e avrà luogo la trascrizione. Il primo nucleotide del neofilamento di RNA trattiene i tre gruppi fosfato mentre quelli successivi vengono privati di due gruppi fosfato attraverso una reazione esoergonica.
Quando, durante la trascrizione, nel DNA si incontreranno particolari sequenze di basi (alla fine del gene) si avrà il termine della trascrizione. Poiché i due filamenti si legano tramite appaiamento delle basi azotate complementari, questi sono tra loro antiparalleli. La direzione di lettura del DNA è 3'->5' mentre quella di trascrizione è 5'->3'. In genere per la trascrizione di un gene si utilizza solo un filamento della doppia elica, mentre l'altro filamento potrà essere utilizzato per la trascrizione di un altro gene.
DNA
Vai a: Navigazione, cerca

Schema della replicazione del DNA
L'acido desossiribonucleico o deossiribonucleico (DNA) è, dal punto di vista della biochimica, un polimero organico i cui monomeri sono i desossiribonucleotidi. È presente in tutti gli organismi viventi. Lo si trova nei cromosomi degli eucarioti, dei procarioti, oltre che nei plasmidi, nei mitocondri e in molti virus. È una molecola molto importante perché trasporta l' informazione genetica necessaria alla trasmissione dei caratteri ereditari. Ogni proteina presente negli organismi viventi deriva da un processo di sintesi che trae origine dall'informazione immagazzinata nel DNA.
Ogni nucleotide è formato da tre parti: una
molecola di desossiribosio (uno zucchero semplice, appartenente ai pentosi), un
gruppo fosfato e una base azotata (citosina, guanina, adenina o timina).
L'atomo di carbonio in
Di solito il DNA è a doppio filamento: è formato da due catene orientate in verso opposto, unite da legami idrogeno tra le basi azotate. Ogni sequenza è determinata dall'altra, in quanto la regola di appaiamento A-T, G-C è imposta dalla dimensione delle basi e dal numero e dalla disposizione dei legami idrogeno che esse possono formare. Si dice anche che i due filamenti sono complementari.
I due filamenti sono avvolti l'uno attorno
all'altro in una doppia elica, struttura che corrisponde ad un minimo di
energia. Quelle di DNA sono molecole molto lunghe: un cromosoma umano medio
contiene un doppio filamento di DNA lungo
REPLICAZIONE
La replicazione del DNA è una reazione di polimerizzazione che ha come reagenti i quattro tipi di desossiribonucleosidi trifosfati (dNTP: dATP, dCTP, dGTP, dTTP). Benché nel filamento venga incorporato solo un fosfato, i nucleotidi di partenza devono essere trifosfati, solo così posseggono infatti l'energia necessaria per la reazione. È necessaria la presenza di un DNA a filamento singolo che funge da stampo, che determina la sequenza del filamento da costruire.
La reazione è catalizzata dalle DNA polimerasi, enzimi capaci di costruire una nuova catena nel verso 5'-3' individuati da Arthur Kornberg nel 1958 tramite un famoso esperimento [1]. Esse non sono in grado di iniziare un filamento ex novo, possono solo allungare un filamento polinucleotidico preesistente. È necessario quindi un innesco. Questo consiste di solito in un breve frammento di RNA appaiato allo stampo, prodotto da una RNA polimerasi detta primasi.
Per iniziare la replicazione, il DNA a doppia elica deve essere parzialmente denaturato da particolari proteine. Queste sono le elicasi, enzimi che separano attivamente i due filamenti usando l'energia dell'ATP, e le proteine denaturanti, o proteine destabilizzatrici dell'elica, non enzimatiche, che possono denaturare il DNA legandosi selettivamente alle porzioni a singolo filamento e stabilizzandole. Queste attività producono una forca replicativa, che migra esponendo progressivamente filamenti non appaiati, che possono essere replicati.
![]()
Forca replicativa
Poiché le polimerasi lavorano solo in senso 5'-3', un filamento (chiamato Filamento a replicazione progressiva) può essere replicato in modo quasi continuo, man mano che viene esposto, l'altro (Filamento a replicazione regressiva) risulta disseminato da brevi filamenti di DNA di nuova sintesi (i frammenti di Okazaki), ognuno dei quali reca all'inizio l'innesco di RNA. I nuovi filamenti devono essere quindi completati mediante la rimozione degli inneschi da parte di endonucleasi e il riempimento degli spazi rimasti ad opera di polimerasi di riparazione. Successivamente tutti questi frammenti di DNA di nuova sintesi del filamento in ritardo vengono legati dalla DNA-ligasi.
Il risultato della replicazione sono due doppie eliche identiche (salvo errori avvenuti durante il processo, che portano alla sa di mutazioni) costituite da un filamento preesistente e uno neoformato: per questa ragione la replicazione si dice semiconservativa.
Nelle molecole di DNA circolari dei Procarioti si ha una sola regione di Origine della replicazione dal quale partono due forche replicative (la struttura prende il nome di bolla di replicazione). Quando le due forche si incontrano dal lato opposto la replicazione è completata.
Negli Eucarioti la replicazione di ogni cromosoma inizia in più punti.

Animazione di un frammento di DNA
Le basi azotate (adenina A, citosina C, guanina G, timina T), possono essere immaginate come le quattro lettere dell'alfabeto delle informazioni genetiche della cellula. Utilizzando gruppi di tre lettere si possono avere fino a 64 combinazioni diverse (43), che vanno a coprire i venti diversi amminoacidi esistenti.
Ad esempio l'adenina ripetuta in una serie di tre ('AAA') rappresenta un particolare amminoacido: la fenilalanina. Poiché esistono 64 triplette possibili e 20 amminoacidi, il codice genetico è degenerato (ridondante), ovvero alcuni amminoacidi possono essere codificati da più triplette diverse, non ci sarà però mai un'ambiguità, ad ogni tripletta corrisponderà un solo amminoacido. Esistono infine triplette che non codificano per amminoacidi ma per codoni di stop, ovvero indicano il punto in cui in un gene termina la parte che codifica per la proteina corrispondente.
Nell'RNA la timina si lega all'uracile (contraddistinto dalla lettera U) che rappresenta la quinta base della complessa struttura dell'elica del DNA/RNA
L'uracile particolare fu la prima base scoperta negli studi sul DNA, per cui introducendo la molecola nel filamento, la cellula produceva timina. Questo sorprese i ricercatori.
CROMOSOMI
In biologia, il cromosoma è un corpuscolo che appare nel nucleo cellulare di una cellula eucariota durante la mitosi. I cromosomi sono spesso presenti in coppie, 23 nella specie umana, di cui 22 coppie sono cromosomi omologhi (cioè simili) detti autosomi ed una coppia di cromosomi diversi che sono i cromosomi sessuali. Tutti i cromosomi sono portatori dei caratteri ereditari. Le cellule che hanno coppie di cromosomi omologhi sono dette diploidi (2n), mentre sono definite aploidi (n) quelle che possiedono solo un cromosoma per tipo.
I nuclei delle cellula eucariotica contengono un materiale che si colora intensamente con certi coloranti istologici e viene quindi detto cromatina. Durante l'interfase la cromatina non rivela alcuna struttura, se non la differenziazione tra una componente maggioritaria più lassa (eucromatina) e una più condensata (eterocromatina).
Durante la divisione cellulare la cromatina si suddivide in un numero ben definito di corpiccioli con la dimensione maggiore dell'ordine del micrometro: i cromosomi. Nella cellula appena formata, i cromosomi hanno forma di bastoncelli: l'unica struttura evidente al microscopio è una strozzatura detta centromero.
Nella metafase i cromosomi hanno una forma a X, dovuta al fatto che si sono quasi completamente duplicati e risultano formati da due cromatidi identici, uniti per il centromero che si divide per ultimo.
Al microscopio ottico, i cromosomi sono distinguibili tra loro per le dimensioni e per la 'forma', ossia per la posizione del centromero. Ulteriori distinzioni si possono effettuare con opportuni trattamenti chimici, che evidenziano un bandeggio: l'alternanza di bande con diversa pigmentazione.
Numero e struttura dei cromosomi costituiscono il cariotipo, ben evidenziabile (e fotografabile) durante la metafase, in cui i cromosomi si dispongono nella piastra metafasica.
La cromatina è costituita permanentemente da DNA e proteine. Il Dna è avvolto attorno a cilindretti formati dai quattro tipi di istoni, proteine basiche: tale fibra fondamentale si chiama cromonema ed è ulteriormente avvolta in strutture di ordine superiore.
MITOSI
Il ciclo cellulare si suddivide in 3 parti: l'interfase, in cui la cellula si prepara alla divisione; la mitosi, periodo di gran lunga più breve in cui la cellula si divide che viene descritto nei particolari in questa ina; il periodo detto G0, più o meno definitivo, in cui la cellula si specializza nella sua funzione e non è in grado o non è stimolata a riprodursi.
Il processo inizia con la condensazione della cromatina che avviene grazie alla presenza di proteine istoniche che fungono da centri primari di organizzazione del riavvolgimento del DNA, primo ordine di spiralizzazione, e della topoisomerasi II che, oltre alla sua funzione catalitica, agisce come centro di organizzazione del secondo ordine di spiralizzazione; poi seguono un terzo ordine di cui non si conoscono le proteine implicate e forse è conseguenza della tensione accumulata dalle precedenti spiralizzazioni; poi questo grosso superfilamento viene prima impaccato formando delle anse che poi si riuniscono formando il cromosoma visibile.
La mitosi si svolge in 4 fasi distinte e susseguenti;
Profase
Metafase
Anafase
Telofase
La durata media di questo meccanismo di riproduzione cellulare" class="text">riproduzione cellulare varia in media, negli organismi superiori, tra le 10 e le 30 ore.
Eventi che precedono la mitosi
Prima della mitosi avviene l'interfase, momento molto importante nella vita della cellula, infatti proprio in questa fase, che si suddivide in G1, S e G2, gli organelli della cellula aumentano e nella fase S il DNA si duplica.
Profase
I 46 cromosomi (nel caso dell' uomo), si
condensano e sono visibili al microscopio ottico sotto forma di doppi
bastoncelli basofili, i cromatidi gemelli, che sono agganciati tra loro in un
punto centrale detto centromero grazie ad un complesso sistema di interazioni
tra il DNA e numerose proteine.
Viene sintetizzato un secondo centrosoma ed entrambi appaiono circondati da una
coltre di microtubuli. Il ciclo si potrebbe interrompere in questo punto se
alla coltura si aggiungesse la tossina falloidina che impedisce la formazione
dei filamenti di microtubuli, questo si fa quando si vogliono visualizzare al
microscopio i cromosomi. L'apparato del Golgi, il reticolo endoplasmatico in
questa fase si scompongono in piccole vescicolette che si distribuiscono
uniformemente in tutto il citoplasma; anche la membrana nucleare, grazie alla
sua struttura a doppia membrana si scompone similmente agli organelli citati
prima.
Metafase
(meta- = successiva)
I due centrosomi si portano ai poli opposti della cellula ed agiscono come centri di organizzazione microtubulare, catalizzando l'allungamento ed assicurando il corretto orientamento dei microtubuli che andranno a breve a legarsi al centromero di uno dei due cromatidi gemelli. In questa fase si possono verificare degli errori e due microtubuli si possono agganciare allo stesso cromatidio dando poi una cellula lia mutilata e non vitale. Questa fase viene chiamata anche prometafase, che significa prima parte della metafase.
Le 46 coppie di cromatidi vengono portate nella parte mediana della cellula, formando la 'piastra equatoriale' in cui un piano immaginario passante per i centromeri divide le coppie di DNA.
Anafase
(ana- = ulteriore)
I cromatidi si dividono in due parti e vengono avvicinati ai poli del fuso centrale, in modo da ripristinare, per ogni polo, il numero originario di cromosomi.
Telofase
(telo- = finale)
I cromosomi si despiralizzano. Intorno ai due nuovi complessi cromosomici ricompaiono le membrane nucleari e gli organelli si ricompongono. La telofase si conclude con la citodieresi, che significa divisione della cellula in due; la cellula si divide al centro formando due cellule lie, esattamente identiche alla cellula madre ma più piccole. Questo avviene grazie ad un anello di actina creatosi al centro della cellula madre che contraendosi stringe la cellula al centro, a questo punto proteine specializzate operano la fusione e la separazione della membrana in punti specifici e le due cellule si separano.
In alcune cellule la telofase non avviene e si accumulano all'interno di uno stesso nucleo di una stessa cellula da due ad alcune decine di corredi cromosomici. Questo tipo di cellule si chiamano plasmodi e l'esempio principe sono i protozoi del genere plasmodium come il plasmodium malariae; anche cellule umane vanno incontro a questo processo o patologicamente, come le cellule tumorali, o fisiologicamente come nel megacariocita e forse (in attesa di una conferma dalla ricerca) in una minoranza di cellule cardiache ed epatiche, anche i macrofagi iperattivati in un granuloma vanno incontro a questa 'mitosi mutilata'.
Raddoppiamento dei cromosomi
Il nucleo della cellula somatica umana possiede 46 cromosomi, dividendosi questo numero diventerebbe 23, perdendo parte del corredo cromosomico. L'unico modo per scindersi e rimanere allo stesso numero è che la divisione avvenga dopo un raddoppiamento, lasciando due copie di ogni singolo cromosoma.Quindi, il requisito essenziale per una cellula che voglia riprodursi, è il raddoppiamento dei propri cromosomi, che equivale a dire lo sdoppiamento del proprio acido desossiribonucleico (DNA).
Nel periodo della mitosi, la cellula è estremamente vulnerabile, basta un piccolo errore nello sdoppiarsi, dovuto al caso o a cause esterne, e le cellule che ne derivano saranno non vitali.
Differenziazione e maturazione
La differenziazione e la maturazione, sono due processi che si oppongono alla mitosi, che la contrastano e tendono a bloccarla. Più una cellula è immatura e più si divide. Inversamente, una cellula matura, specializzata, ha una ridotta capacità proliferativa.
La meiosi è il meccanismo di divisione mediante il quale una cellula eucariotica diploide da origine a quattro cellule aploidi. È molto simile alla mitosi.
Si compone principalmente di due parti:
La precede un normale ciclo cellulare.
Durante
[modifica]
La seconda divisione meiotica è identica alla mitosi, solo che genera due cellule aploidi, perché non è preceduta da un ciclo cellulare adeguatamente fornito di fase S, e quindi avviene in presenza di un patrimonio genetico 2N invece che 4N (e da origine quindi a cellule N invece che 2N)
PRIMA DIVISIONE MEIOTICA
In sintesi, nella prima divisione meiotica si evidenziano i cromosomi, ciascuno costituito da due cromatidi. Questi cromosomi (metà di origine paterna e metà di origine materna), dopo aver subito alcuni processi durante la profase (in particolare il crossing-over, di cui parleremo successivamente), si portano al piano equatoriale della cellula. Qui, senza dividersi nei due cromatidi, si attaccano alle fibre del fuso per migrare verso i due poli in modo tale che, di ogni coppia di cromosomi omologhi, una si dirige verso un polo e l'altra al polo opposto. A conclusione della prima divisione meiotica, si hanno così due cellule, ciascuna con la metà esatta dei cromosomi omologhi.
SECONDA DIVISIONE MEIOTICA
La seconda divisione meiotica non è preceduta da alcuna duplicazione del DNA. I cromosomi, costituiti da due cromatidi, si portano all'equatore e si attaccano alle fibre del fuso; i due cromatidi di ciascun cromosoma si separano migrando ai poli. Si formano così quattro cellule, ciascuna con un corredo aploide di cromosomi e con un diverso assortimento dei cromosomi di origine materna e paterna. Durante questa separazione vi è una distribuzione indipendente dei cromosomi paterni e materni per cui, alla fine, vi sarà un diverso assortimento dei cromosomi nelle quattro cellule lie.
Da un punto di vista genetico, la meiosi assume una grande importanza perché rappresenta il modo in cui possono formarsi nuove combinazioni di geni e, quindi, rende possibile la variabilità genetica tra individui della stessa specie. Infatti, già con il crossing-over, ovvero con lo scambio di porzioni di DNA tra cromatidi di due cromosomi omologhi, al momento della profase I, avviene una prima modificazione dell' assortimento di geni rispetto a quello della cellula madre. Inoltre, occorre considerare che la divisione dei due cromosomi omologhi durante la fase di anafase I avviene in modo casuale: ciò significa che non è prestabilito il polo della cellula verso cui migrerà ciascun cromosoma. Dunque, a partire da una cellula madre, si formano con la prima divisione meiotica due cellule aploidi che sono geneticamente differenti tra loro e diverse da qualsiasi altra coppia di cellule che derivano dalla stessa cellula madre. La variabilità genetica, assicurata anche dai meccanismi di mutazione spontanea, assume un ruolo essenziale nei processi evolutivi, secondo il concetto di selezione naturale.
 |  |
|
Privacy
|
© ePerTutti.com : tutti i diritti riservati
:::::
Condizioni Generali - Invia - Contatta
