|
|
| biologia |
|
|||||
|
DNA & Cromosomi
La vita dipende dalla capacità cellula - STRUTTURA DELLE CELLULE EUCARIOTE" class="text">delle cellule di conservare, estrarre e tradurre le istruzioni genetiche richieste per creare e mantenere un organismo vivente. Questa informazione ereditaria e trasmessa da una cellula alle cellule lie durante la divisione cellulare e da una generazione di un organismo alla successiva attraverso le cellule riproduttive.
Queste istruzioni sono conservate in ogni cellula vivente come geni, elementi che contengono informazioni che determinano le caratteristiche di una specie nel suo insieme e degli individui che la compongono. I primi cercatori cominciarono a domandarsi quale fosse la Struttura chimica di tali geni. Agli inizi degli anni 40 cominciarono ad emergere a e risposte. Infatti alcuni ricercatori, proprio in questo periodo, scoprirono da studi semplici su funghi, che l'informazione genetica consiste primariamente in istruzioni per produrre proteine. Quest'ultime sono macromolecole che svolgono la maggior parte delle funzioni cellulari: servono da unità da costruzione per le strutture cellulari informano gli enzimi che catalizzano tutte le reazioni chimiche della cellula, regolano l'espressione dei geni e rendono le cellule capace di muoversi e di comunicare tra loro.
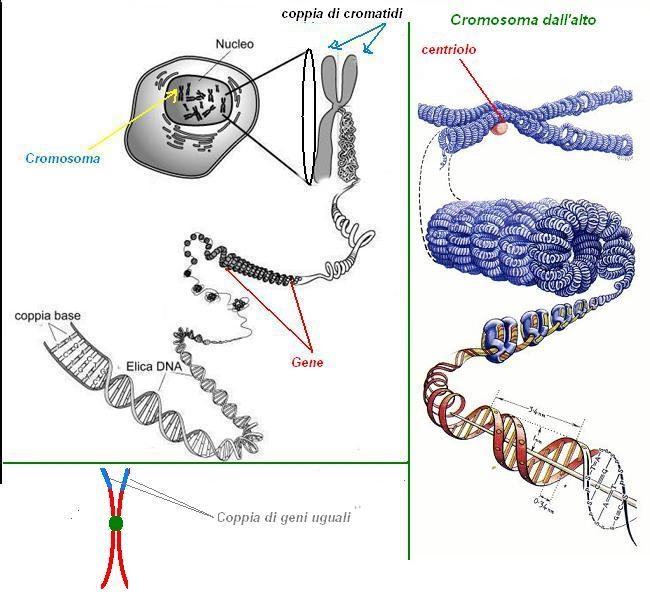
I biologi, inoltre, avevano riconosciuto che i geni erano portati su cromosomi scoperti nel diciannovesimo secolo come strutture filamentose del nucleo di una cellula eucariota che diventavano visibili quando la cellula cominciava a dividersi.
Ma all'avanzamento cruciale fatto negli anni ' 40 è stato l'identificazione dell' acido desossiribonucleico (Dna) come trasportatore dell'informazione genetica. Inoltre la maggior parte dei ricercatori riteneva che il materiale genetico fosse costituito dalle proteine e che il Dna, essendo formato da soli quattro nucleotidi, fosse troppo semplice per giustificare la varietà e la ricchezza dell'informazione genetica. Queste ipotesi però si rivelò sbagliata. Infatti la prova decisiva fu fornita nel 1952 da Hershey e Chase i quali dimostrarono che l'informazione genetica era contenuta nel Dna. Ciò fu dimostrato suoi batteriofagi, i quali contengono un unico tipo di acido nucleico (Dna o RNA) che porta solo l'informazione genetica relativa alla sintesi dei propri elementi costitutivi. Essi, inoltre, per riprodursi devono iniettare in cellule ospiti il loro materiale ereditario, contenuto in una molecola di Dna.
Nel frattempo rimanevano ancora oscure le modalità con cui veniva trasmessa l'informazione ereditaria da una cellula all'altra e il modo in cui le proteine specificavano le istruzioni nel DNA.
All'inizio degli anni '50 Rosalind Franklin iniziò ad applicare le sue competenze a proposito delle tecniche di diffrazione raggi X per cercare di determinare la struttura atomica tridimensionale di una molecola di Dna. I primi risultati indicarono che il Dna era composto da due filamenti del polimero avvolti in un'elica.
In questo modo noto che vi erano due forme di DNA. Infatti ad alta umidità (quando bagnato) il Dna diventa una fibra lunga e sottile, mentre quando invece asciutto, il Dna è una fibra corta e grossa. Queste Dna vennero definiti rispettivamente ' B' e 'A'. Il lavoro sul Dna fu successivamente diviso tra Franklin e Wilkins: il primo analizzò la forma 'A', mentre secondo la 'B'. Entro la fine del 1951 fu accettata che la forma 'B' del DNA era una spirale, ma si dubitava fortemente che la seconda forma avesse una struttura elica. Nel corso del 1952 fu applicata la funzione di Patterson alle foto dei raggi x di Dna che avevano realizzato precedentemente. Il lavoro fu difficile, ma delle immagini ottenute, tramite la diffrazione a raggi x, Franklin notò che nella fotografia 51 era rappresentata la forma B del DNA.
Nel 1951 Watson e Crick avviarono un'intensa collaborazione intellettuale che portò alla risoluzione della struttura del DNA in non meno di un anno e mezzo. Ma ciò sarebbe stato realizzato con un circa un anno di anticipo da Pauling grazie alla sua lunga esperienza, ma ciò non avvenne a causa del divieto di visitare l'Inghilterra per ragioni politiche. In questo modo non poté avere accesso ai dati sulla diffrazione raggi X.
Nel corso del 1952, in ogni caso, a Watson e Crick fu richiesto di non lavorare sui modelli molecolari della struttura di DNA, ma l'attività ufficiale dei due sarebbe stata l'analisi con diffrazione raggi X del virus del mosaico del tabacco. Nel 1954, Boston scoprirà dalla diffrazione raggi X che la struttura del virus è di tipo elicoidale. Nonostante ciò Watson è cricco continuarono a lavorare sulla struttura del Dna, anche se in seguito furono costretti a rivolgersi alla Franklin, la quale aveva già portata a termine notevoli lavori sul Dna e poteva fornire loro numerose conoscenze chiave per portare a termine il lavoro. Servendosi dei lavori non ancora pubblicati della Franklin e di Wilkins, i due poterono dedurre la struttura a doppia elica che pubblicarono nel 1953 sulla rivista "Nature". A disegnare il bozzetto fu Speed, pittrice e moglie di Crick. In particolare la Franklin contribuì nell'individuare la funzione dei gruppi fosfato degli acidi nucleici. La ricercatrice chiarì la loro posizione esterna, suggerendone la funzione di supporto. Watson e Crick ne conclusero dunque che l'informazione risiedesse nell'ordine delle quattro basi azotate.
Nel 1962, dopo la morte di Franklin (a causa di un tumore provocato, probabilmente, dalle alti dosi di raggi X a cui si era esposto nel corso dei suoi esperimenti), Watson e Crick ricevettero il Premio Nobel per la medicina.
v Struttura e funzione del DNA:
Dal punto di vista chimico, il Dna è un polimero lineare o organico costituito da monomeri chiamati nucleotidi. I nucleotidi sono costituiti da uno zucchero cinque atomi di carbonio al quale sono legati uno più gruppi fosfato e una base che contiene azoto. I costituenti principali del nucleotidi, quindi, sono:
Un gruppo fosfato
Il zucchero pentoso
Una base azotata.
I nucleotidi sono uniti covalentemente fra loro in una catena tramite gli zuccheri e i fosfati, che formano così una "ossatura" di zucchero-fosfato-zucchero-fosfato, alternati

![]()
![]()
![]()
![]()
![]() Zucchero fosfato
Zucchero fosfato
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]() base
base
G G zucchero Fosfato
![]()
nucleotide
Le basi azotate utilizzate nella formazione dei nucleotidi del DNA sono quattro:
Adenina (A);
Guanina (G);
Citosina (C);
Timina (T).
Una molecola di Dna consiste di due lunghe catene polinucleotidiche composta dai quattro tipi di nucleotidi ( o subunità: A, G, C, T). Ciascuna di queste catene è nota come catena di DNA o filamento di Dna. I due filamenti di DNA tenuti insieme da legami idrogeno tra le basi appaiate.
Nel caso del DNA, invece, i nucleotidi hanno come zucchero, il deossiribosio legato ad un singolo un gruppo fosfato ( da cui deriva appunto, il nome acido deossiribonucleico) e una base azotata (A, C, G e T).
La struttura laterale delle DNA e composta da unità ripetute ed alternate di gruppi fosfato e di 2 deossiribosio, uno zucchero pentoso (a cinque atomi di carbonio) che si lega ai fosfati adiacenti attraverso legami fosfodiesterici. Quest'ultimo è un legame covalente (ovvero una condivisione di elettroni che mantiene uniti, con legame forte, due atomi di una molecola che si forma in una catena polinucleotidica (come il DNA o l'RNA) tra il fosfato associato al carbonio in posizione 5 dello zucchero di un nucleotide e il carbonio in posizione 3 dello zucchero del nucleotide successivo.
Il polinucleotide che si forma grazie a questi legami ha una direzionalità intrinseca: ad una estremità della catena il primo nucleotide presenta un ossidrile libero OH) al carbonio in posizione 3 dello zucchero, mentre all'atra estremità l'ultimo nucleotide presenta un gruppo fosforico PO4) al carbonio in posizione 5 dello zucchero Le due diverse estremità, che indicano la polarità, vengono dette estremità 3' (-OH) ed estremità 5' (-PO4) ovvero estremità tre primo e cinque primo.
In una doppia elica, il senso di un filamento e opposta a quella del filamento complementare. I due filamenti, quindi, che costituiscono una doppia elica sono detti "antiparalleli , cioè la polarità di un filamento è orientata in senso opposto a quella dell'altro filamento. Una conseguenza di questi requisiti è che ciascun filamento di una molecola di DNA contiene una sequenza di nucleotidi esattamente complementare alla sequenza nucleotidica dell'altro filamento.
I due filamenti polinucleotidici sono tenuti insieme nella doppia elica del DNA da legami idrogeno tra le basi dei due filamenti opposti: tutte le basi sono rivolte verso l'interno dell'elica, mentre l'ossatura zucchero-fosfato rimane all'esterno.
La struttura a doppia elica del Dna è stabilizzata dai legami idrogeno che si instaurano tra le basi presenti sui due filamenti. Le quattro basi che sono state individuate nelle Dna sono l'adenina (A), la citosina (C), la guanina (G) e la timina (T). Adenina e guanina sono composti eterociclici chiamati purine, mentre citosina e timina sono anelli piramidinici (piramidine). Di quest'ultimo tipo esiste una quinta base chiamata uracile (U), ma essa non è di norma presente nelle catene di Dna. Infatti essa si trova sui filamenti di RNA al posto della timina, essa determina la differenza tra DNA ed RNA, appunto per la mancanza di un gruppo metile. L'uracile è presente nel DNA solo come il prodotto della degradazione della citosina. Al contrario, è molto più frequente individuare la timina all'interno di molecole di RNA, a causa della metilazione enzimatica di diversi uracili.
Piramidine: ad anello




PURINE doppio anello



La differenza sostanziale riguarda lo zucchero. Sebbene sia nel DNA che nell'RNA sia presente uno zucchero pentoso, nell'RNA (acido ribonucleico) è presente il ribosio, che ha un gruppo ossidrilico in posizione 2 dello zucchero, assente nel desossiribosio, lo zucchero pentoso caratteristico del DNA (acido desossiribonucleico). Questa caratteristica espone l'RNA ad un'idrolisi più rapida, rendendolo un acido nucleico non idoneo alla conservazione dell'informazione.
v Regole di chargaff :
Fu Chargaff a scoprire i particolarità del legame e dei rapporti che si creavano tra le quattro basi azotate del DNA. In base ai suoi studi poté distinguere due diverse:
PURINE = PIRAMIDINE
La prima regola mostra l'esistenza di un rapporto 1:1 tra le purine (A+ G) e le piramidine (T+ C) contenute nel Dna di una cellula. Il rapporto è costante in tutte le specie. Questo rispecchia la diversità genetica delle diverse specie.
![]()
La seconda regola mostra che in una molecola di Dna doppio filamento la percentuale di adenina eguaglia quella di timina; e la concentrazione di citosina quella della guanina (%A=%T; %C=%G). Questo vale per il Dna estratto dalle cellule di tutti gli organismi, anche se considerando organismi diversi Le percentuale avranno valori diversi. Questa semplice regola è stata uno degli elementi essenziali che hanno permesso la formulazione del modello di DNA da parte di Watson e Crick. (cioè la quantità di guanina è pari a quella della citosina, mentre la quantità di adenina è pari a quella della timina).
![]()
v Appaiamento complementare delle basi
Ogni tipo di base presente su un filamento forma un legame con la base posta sul filamento opposto. Tale evento è noto come appaiamento complementare delle basi . Le basi purine formano legami idrogeno con le basi piramidine: A può legare solo con T; G può legare il solo con C. L'associazione di due basi viene comunemente chiamata "paio di basi". Dal momento che legami idrogeno non sono covalenti, i due filamenti possono essere rotti e riuniti in modo relativamente semplice. Praticamente i due filamenti possono essere allontanati tra loro come avviene per una cerniera, sia dalle alte temperature che da un'azione meccanica (come avviene durante la replicazione del DNA). Conseguenza di questa complementarietà è che tutte le informazioni contenute nella doppia elica possono essere duplicate a partire da entrambi filamenti.
I due tipi di paia di basi formano un numero differente di legame idrogeno:
A e T ne formano due.
G e C ne formano tre.
Per tale motivo, la stabilità del legame GC è decisamente maggiore di quello AT. Di conseguenza, la stabilità complessiva di una molecola di Dna è direttamente correlata alla frequenza di GC presenti nella molecola stessa, nonché dalla lunghezza delle elica: una molecola di Dna è dunque tanto più stabile quanto più contiene GC e quanto più è lunga.
Oltre alla complementarità un'altra caratteristica del DNA è la anche antiparallelismo. Infatti i due filamenti appaiati sono orientati in senso opposto (5'-3') e (3'-5').
Ereditarietà:
I geni portano l'informazione biologica che deve essere copiata accuratamente per essere trasmessa alla generazione successiva ogni volta che una cellula si divide formando due cellule lie. Il Dna codifica l'informazione tramite l'ordine, o sequenza, dei nucleotidi in ciascun filamento. Ciascuna base A,T,G,C può essere considerata come una lettera con cui sono scritti messaggi biologici. L'informazione genetica conservata nel Dna di un organismo contiene le istruzioni per tutte le proteine che l'organismo dovrà sintetizzare.
La serie completa di informazioni nel Dna di un organismo si chiama genoma e porta le informazioni per tutte le proteine che l'organismo dovrà sintetizzare. A ciascuna divisione cellulare, la cellula deve copiare il suo genoma per passarlo ad entrambe le cellule lie. La scoperta della struttura del Dna ha anche rilevato il principio che rende possibile questa copiatura: poiché ciascun filamento di Dna contiene una sequenza di nucleotidi che è esattamente complementare alla sequenza nucleotidica del filamento partner, ciascun filamento può agire da stampo, o forma, per la sintesi di un nuovo filamento complementare. In altre parole, se designiamo due filamenti di Dna come S e SI, il filamento S può servire da stampo per un nuovo filamento SI, mentre il filamento SI può servire da stampo per formare un filamento S.
La capacità di ciascun filamento di una molecola di Dna di agire da stampo per produrre un filamento complementare rende una cellula in grado di poter copiare, o replicare, i suoi geni prima di passarli ai suoi discendenti.
Il Dna negli eucarioti:
Quasi tutto il Dna in una cellula eucariota è confinato nel nucleo, che occupa circa il 10% del volume cellulare totale. Questo timento è delimitato da un involucro nucleare formato da due doppi strati lipidici concentrici, perforati ad intervalli regolari da grandi pori nucleari che trasportano molecole fra il nucleo e il citosol.
La compattazione del nucleo è un principio importante della biologia. Infatti esso facilita le reazioni biochimiche a causa dell'alta concentrazione sia dei substrati che degli enzimi che agiscono su di essi.
Il Dna cromosomico e il suo compattamento nella fibra di cromatina
La funzione più importante del DNA è quella di portare i geni, i quali contengono sia l'informazione che specifica tutte le proteine che costituiscono un organismo che l'informazione che indica quando, in quale tipo di cellula e in quale quantità ciascuna proteina deve essere prodotta. Ciascuna cellula umana contiene, se esso viene steso, approssimativamente 2 metri di DNA. Eppure il nucleo di una cellula umana che contiene il Dna, ha un diametro di soli 6 ηm. Il compito più complesso risulta proprio quello di compattare il DNA. Tale compito è svolto da proteine specializzate che legano il DNA e lo ripiegano, generando una serie di avvolgimenti e ansie che forniscono livelli sempre più alti di organizzazione, impedendo così che il DNA diventi un groviglio impossibile da maneggiare.
I cromosomi:
Negli eucarioti il Dna umano è diviso in una serie di cromosomi diversi. Il genoma umano, per esempio, è distribuito su 24 cromosomi diversi. Ciascuno cromosoma consiste di una singola molecola di Dna estremamente lunga e associata proteine che ripiegano e compattano il sottile filamento di Dna in una struttura più compatta. E complesso costituito da Dna e proteine si chiama cromatina (dal greco o croma "colorata"). Ma i cromosomi sono anche associati a molte proteine necessarie per i processi di espressione genica, di replicazione del DNA e della riparazione del DNA stesso.
I batteri, invece, portano i loro geni su una singola molecola di Dna, che di solito ha una forma circolare. Tale Dna è anch'esso associato proteine che lo compattano e lo condensano, ma tali proteine sono differenti da quelle che svolgono le stesse funzioni negli organismi eucarioti. Sebbene anche quello batterico sia definito "cromosoma" non ha la stessa struttura dei cromosomi eucariotici e poco si sa del modo in cui esso sia compattato.
Con l'eccezione delle cellule germinali e pochi tipi cellulari che non possono moltiplicarsi e sono completamente privi di DNA (come ad esempio i globuli rossi) ciascuna cellula umana contiene due coppie di ciascuno cromosoma, una ereditata dalla madre e una dal padre. I cromosomi materno e paterno di una coppia sono chiamati cromosomi omologhi.
L'unica coppia di cromosomi non omologhi è composta dai cromosomi sessuali nei maschi, in cui un cromosoma Y è ereditato dal padre e un cromosoma X dalla madre (nelle donne invece la coppia è ereditata completamente dalla madre XX).
Così, ciascuna cellula umana contiene un totale di 46 cromosomi, di cui 22 coppie sono comuni a maschi più due cromosomi sessuali ( XY nei maschi e XX nelle femmine ). L'insieme dei 46 cromosomi umani della mitosi si chiama cariotipo umano. È possibile sapere se parti di cromosomi sono stati perse o scambiate fra cromosomi, grazie all'ibridazione del DNA che permette di distinguere i cromosomi umani "dipingendoli" di colori diversi. Eventuali cambiamenti possono essere rilevati da variazioni della colorazione dei cromosomi.
La funzione più importante dei cromosomi è quella di portare i geni, ovvero le unità funzionali dell'eredità. Un genere, di solito è definito come una segmento di Dna che contiene le istruzioni per produrre una particolare proteina. Ma una discreta percentuale di geni produce, invece di una proteina, una molecola di RNA che, come le proteine, svolge una serie di diversificata di funzioni strutturali e catalitiche nella cellula.
I cromosomi di molti organismi eucarioti contengono, oltre ai geni, una grande quantità di Dna in eccesso che non sembra portare informazioni cruciali. Tale DNA è definito Dna spazzatura in modo tale da indicare che la sua utilità non è stata ancora dimostrata.
La differenza totale di geni varia da un organismo all'altro. Approssimativamente esso varia dai 500 geni per i batteri semplici fino ai 30.000 per l'uomo. È pero da tener conto che più è complesso un organismo e più è grande il suo genoma, ma a causa del DNA in eccesso la relazione non è sistematica. A causa di differenze nella quantità di DNA in eccesso, i genomi di alcuni organismi simili possono variare di centinaia di volte nel loro contenuto di DNA, pur avendo più o meno lo stesso numero di geni. Analogamente la divisione del genoma in cromosomi differisce anch'essa da una specie eucariotica all'altra. Da ciò si deduce che non c'è una relazione semplice fra numero di cromosomi, complessità della specie e dimensioni totali del genoma.
Disposizione dei geni sul cromosoma:
La disposizione dei geni sul cromosoma fu scoperta nel 1999 analizzando la sequenza di uno dei cromosomi umani più piccoli: il cromosoma 22. Tale cromosoma corrisponde approssimativamente all'1,5% dell'intero genoma umano. Tale cromosoma è costituito da 3,2 x 109 coppie di nucleotidi ed è distribuito in 22 autosomi e due cromosomi sessuali. Con il termine "genoma umano" ci si riferisce alla sequenza di nucleotidi completa del DNA dei 24 cromosomi. Essendo diploide, una cellula somatica romana contiene circa il doppio di questa quantità di DNA. Gli esseri umani differiscono l'uno dall'altro in media di un nucleotide ogni 1000.
Quando la pubblicazione della "prima stesura" del genoma umano avvenuta nel 2001, il panorama genetico di tutti i cromosomi umani venne improvvisamente messo a fuoco.
Il primo aspetto sorprendente di questi studi è che è soltanto una piccola percentuale codifichi effettivamente per proteine o RNA strutturali e catalitici. Molto del restante Dna cromosomico è composto da brevi pezzi mobili di DNA che si sono gradualmente inseriti nel cromosoma durante l'evoluzione. Tali elementi sono noti come "elementi trasponibili".
Un secondo aspetto sorprendente, rilevato dagli studi, è rappresentato dalle grandi dimensioni medie dei geni, costituiti da circa 27.000 nucleotidi. Un gene tipico porta nella sua sequenza lineare di nucleotidi l'informazione per la sequenza lineare degli amminoacidi di una proteina. Per codificare una proteina sono necessari, mediamente, soltanto 1300 coppie di nucleotidi. La parte restante del DNA, che costituisce la maggior parte del gene, non è codificante. Tali sequenze sono chiamate introni, mentre le sequenze codificanti sono chiamate esoni. La maggioranza dei geni umani consiste, quindi, in una lunga fila di esoni e introni alternati tra loro, anche se comunque la maggior parte del gene resta costituita dagli introni. La maggioranza dei geni degli organismi con genomi compatti è, invece, priva di introni.
Oltre ad introni ed esoni ogni gene è associato a sequenze regolatrici di Dna che sono responsabili dell'espressione al livello e al momento appropriato e nel tipo appropriato di cellula. Infine, la sequenza nucleotidica del genoma umano ha rilevato che le informazioni cruciali sembrano essere in uno stato allarmante di disordine.
Ulteriori studi hanno rilevato che molte sequenze nucleotidiche dei cromosomi umani sono probabilmente prive di importanza e che gli esoni "galleggiano" in un mare di Dna la cui sequenza nucleotidica ha poca importanza. Questa disposizione rende difficile identificare gli esoni, l'inizio e la fine di un gene e stabilire quanti esoni contiene un gene. Per stabilire ciò gli studiosi si basano sull'osservazione delle sequenze nucleotidiche, notando se la loro funzione si era conservata nel corso dell'evoluzione o se invece a sequenza nucleotidica era mutate casualmente, quest'ultime sequenze sono pive di funzioni. La strategia è perciò quella di confrontare la sequenza umana con quelle regioni corrispondenti di un genoma correlato, in modo da poter notare eventuali mutazioni. Nel caso dell'uomo, ad esempio, viene confrontato con il genoma del topo, in quanto si ritiene che l'uomo e il topo si siano separati da un progenitore comune migliaia di anni fa. Sequenze con regioni simili sono note come regioni conservate. Tali regioni rappresentano esoni funzionalmente importanti e sequenze regolatrici. Le regioni non conservative rappresentano invece Dna la cui sequenza non è generalmente cruciale per la funzione.
Studi ativi di questo tipo hanno mostrato non solo i topi gli esseri umani hanno in comune gli stessi geni, ma anche che grossi blocchi dei genomi di uomo e di topo contengono questi geni nello stesso ordine, una caratteristica chiamata sintonia conservata.
v Formazione e costituzione dei cromosomi
Per formare un cromosoma funzionale una molecola di Dna deve essere capace, oltre che portare i geni, di replicarsi e le copie replicate devono essere separate e divise in modo affidabile nelle cellule lie a ciascuna divisione cellulare. Questo processo avviene in una serie di passi notò come ciclo cellulare. In due fasi di questo ciclo sono coinvolti principalmente i cromosomi; ovvero nell'interfase in cui i cromosomi vengono replicati e nella mitosi dove vengono condensati per poi essere separati e distribuiti ai due nuclei li. Tali cromosomi sono noti cupe "cromosomi mitotici" e sono visualizzati facilmente, mentre nelle altre fasi del ciclo cellulare i cromosomi sono estesi e molta della loro cromatina è sotto forma di lunghi filamenti noti come cromosomi interfasici.
Un cromosoma deve essere capace di replicarsi e le sue copie devono avere le stesse capacità in modo tale che successivamente possano dividersi anch'esse. Queste funzioni basilari sono controllate da tre tipi di sequenze nucleotidiche specializzate nel DNA. Tali sequenze sono state scoperte con esperimenti sui lieviti. Da ciò si è potuto notare che esiste una sequenza nucleotidica che agisce da origine della replicazione del DNA; una sequenza specializzata nota come centromero che permette a ciascun cromosoma duplicato di passare in una cellula lia grazie anche all'aiuto di un complesso proteico noto come cineticore che si forma sul centromero e permette la separazione; una terza sequenza specializzata di Dna forma i telomeri, ovvero l'estremità di un cromosoma.
Centromero![]()
![]()
![]()

Cromatidi
Cromosomi eucariotici:
Le proteine che si legano al Dna per formare i cromosomi eucarioti sono tradizionalmente divisi in due classi generali:
Istoni;
Proteine non istoniche.
Il complesso formato da una di questa classe di proteine e il Dna è nota come cromatina. Gli istoni, scoperti nel 1974, sono le proteine più abbondanti nella cromatina. La loro funzione è quella di avvolgere e compattare lunghissimi filamenti di Dna in modo da poterli contenere nello stretto spazio nucleare. Il Dna si avvolge attorno a gruppi di otto istoni, ottamero di istoni, formando nucleosomi le unità base della cromatina.
La formazione di nucleo atomico avverte una molecola di Dna e in una fila di cromatina avente un aspetto "a collana di perle" lunga circa un terzo della lunghezza iniziale. Fra i due nucleosomi di questo filamento, si trova un'altra molecola di istone che serve ad avvicinare i due nucleosomi successivi, compattando la struttura. La fibra risultante ha un diametro di 30 ηm e costituisce il primo livello di compattamento del DNA.
Ulteriori studi, mediante la cristallografia a raggi X, su una particella centrale di un nucleosoma ne hanno rivelato, nel 1997, la struttura. Un nucleosoma è costituito da un nucleo di istoni a forma di disco intorno al quale è strettamente avvolto il DNA. Una caratteristica comune ai 4 istoni che compongono il nucleo del nucleosoma è il ripiegamento istonico, una struttura formata da tre a-eliche sconnesse da due anse. Oltre a questa caratteristica, ciascun istone del nucleo a una "coda" N-terminale che si estende fuori dal nucleo DNA-istoni. Quest'ultimi, inoltre, sono fra le proteine eucariotiche più conservatrici. Infatti anche un cambiamento piccoli in qualunque posizione sarebbe deleterio per la cellula.
La disposizione dei nucleosomi nel Dna è determinata da due fattori principali. Il primo è la difficoltà di piegare la doppia elica del Dna in due stretti giri all'esterno dell'ottamero di istoni; il secondo, e il più importante, fattore è la presenza di proteine legate al Dna che possono favorire o ostacolare la formazione di un nucleosoma nelle loro vicinanze.
Per molti anni biologi hanno pensato che un nucleosoma rimanesse fisso a causa della stretta associazione fra gli istoni del nucleo e il DNA. Ma recentemente si è scoperto che le cellule eucariote contengono complessi di rimodellamento della cromatina, ovvero macchine proteiche che usano l'energia di idrolisi dell'ATP per cambiare temporaneamente la struttura dei nucleosomi in modo che il Dna sia legato meno strettamente al nucleo di istoni.
La struttura globale dei cromosomi
La maggior parte dei cromosomi nelle cellule interfasiche sono troppo secchi e aggrovigliati per essere visualizzati. In pochi casi, però, si può vedere che i cromosomi interfasici hanno una struttura di ordine superiore precisamente definita e si ritiene che tali caratteristiche siano comuni a tutti i cromosomi interfasici. I cromosomi appaiati nella meiosi chiaramente visibili al microscopio ottico, tramite quest'ultimo si può osservare la loro organizzazione in una serie di grandi anse di cromatina. Tali cromosomi mitotici sono chiamati cromosomi a spazzola.
Alcune cellule di insetto hanno cromosomi interfasici facilmente visibili. Per esempio, molte cellule di alcune larve di musica diventano di dimensioni enormi tramite cicli multipli di sintesi di DNA senza divisione cellulare. Le cellule giganti che ne risultano contengono più del numero di cromosomi standard e per questo sono detti poliploidi ( cellule contenenti più del numero dei cromosomi standard ).
Studi al microscopio ottico negli anni 30 hanno rilevato caratteristiche dei cromosomi interfasici comuni in una grande quantità di organismi. In base a tali studi si sono distinti due tipi di cromatina presenti nei nuclei interfasici delle cellule di organismi eucarioti:
Eterocromatina: cromatina altamente condensata ricca di sequenze ripetute di DNA. Essa non viene trascritta perché troppo compatte inaccessibili all'Rna polimerasi.
Eucromatina: cromatina, rimanente, meno condensata.
La maggior parte del DNA che ripiegato in etero cromatina non contiene geni. Ciò non significa che essa sia inutile, in quanto è responsabile del corretto funzionamento dei telomeri e del centromeri. Inoltre alcuni geni per essere espressi devono trovarsi in regioni di eterocromatina. Tale termine comprende parecchi tipi distinti di strutture di cromatina che hanno come caratteristica comune l'alto grado di organizzazione.
Quando un gene che normalmente è espresso in eucromatina viene sperimentalmente riposizionato in una regione di eterocromatina, questo cessa di essere espresso e si dice che il gene e silenziato. Da ciò si dedotto che l'attività di un gene dipende dalla sua posizione sul cromosoma. Molti effetti di posizione esibiscono un ulteriore caratteristica chiamata da variegazione dell'effetto di posizione. Questi effetti possono derivare da zone di cellule di un gene silenziato è stato riattivato; una volta riattivato il gene è ereditato in questa forma nelle cellule lie. Lo studio della variegazione dell'effetto di posizione ha rilevato due caratteristiche importanti dell'eterocromatina. Per prima cosa il eterocromatina è dinamica; infatti può "diffondersi" in una regione e successivamente "ritirarsi" da essa. Secondo, lo stato di cromatina, sia eucromatina che eterocromatina, tende ad essere ereditato nelle progenie di una cellula.
Le estremità dei cromosomi compattati in eterocromatina forniscono parecchi vantaggi alle cellule in quanto ai a proteggere le estremità dei cromosomi dall'essere riconosciute come cromosomi rotti da parte del macchinario riparatore, aiutano a regolare la lunghezza dei telomeri e possono essere d'aiuto nell'appaiamento accurato e nella segregazione dei cromosomi durante la mitosi.
L'eterocromatina si osserva anche intorno a i centromeri. In molti organismi complessi, compreso l'uomo, ciascun centromero sembra immerso in un tratto molto grande e di eterocromatina che persiste per tutta l'interfase. La struttura e le proprietà di questa, così detta, cromatina centrica non sono ben comprese, ma come le altre forme di eterocromatina, essa silenzia l'espressione di geni. Questa eterocromatina, oltre agli istoni contiene parecchie altre proteine strutturali che compattano i nucleosomi in disposizioni particolarmente dense.
La struttura del Dna A e B:
DNA B: conurazione più frequente 10 paia di basi per giro d'elica, una ogni 3.4 nm. Solco maggiore molto ampio
DNA A: conurazione alternativa compatta della forma B 11 paia di basi per giro d'elica 2.3 nm - solco maggiore ridotto
Proprietà fisiche del DNA:
Viscosità: tutte le volte che si rompe nugolo di una cellula e abbiamo il Dna libero, questo appare il viscoso e non si può prelevare tramite "profeta".
Sedimentazione: se si pone il DNA in ambiente salino (cloruro di sodio) troviamo il Dna tutto raccolto in un punto della soluzione dove la densità propria è uguale a quella delle soluzioni.
Densità: ogni specie di Dna ha una propria densità.
Denaturazione: apertura delle due eliche. Una volta aperte ciascuna delle due eliche funge da stampo.
Il Dna può essere denaturato fornendogli calore. Quando arriviamo ad una temperatura di circa 80 °C abbiamo l'effetto ipercromico
Se forniamo
calure alle due eliche esse hanno un effetto denaturato, cioè si separano ad una determinata
temperatura (Tm). La rinaturazione del DNA porta alla ricostituzione della
doppia elica. Tale effetto è appunto l'effetto Ipercromico. Tm = punto in cui si divide il filamento. Circa 80°C

|
Privacy
|
© ePerTutti.com : tutti i diritti riservati
:::::
Condizioni Generali - Invia - Contatta